Rottnest Island
Located some 19 kilometres off the coast of Western Australia, and visible from the taller buildings of its capital, Perth, lies the Island of Rottnest.Rottnest is WA's best-kept secret. A magic holiday island, with no private vehicles or private land ownership, it is a place tacitly kept for the local people; the increasing number of overseas visitors touring the rest of the State's attractions are subtly not encouraged. Travel around the island is by bicycle, whether you are young or old. Jet-setters would be appalled at the salt-water showers.
Rottnest is an island with a history. It was probably the first landfall of Europeans in the southwest of Australia, when a Dutch party under Willem de Vlamingh landed there in 1696. Vlamingh saw the hordes of the small marsupials called quokkas on the island, and thought that they were large rats; hence the name Rottnest, meaning Rat's Nest in his native language.
Vlamingh was enchanted with the island, too, reporting that "nature had spared nothing to render it delightful -- a terrestrial paradise". At that time, Rottnest was covered with trees, many of which, alas, were cleared away when later settlers attempted to farm the island. These trees and this island have features which, as we will see, have relevance for the ideas expressed in this book.
If there is a family of trees which is typical of Australia, it is the Eucalypts, the Australian gum trees. They exist all over the island continent. But Rottnest has no native gum trees, and moreover, according to Somerville it never did have [75].
Its principal trees are in the genera Melaleuca (Tea Trees) and Callitris (Australian Pines). The Melaleuca species, M. pubescens, is the very distinctive Rottnest Island Tea Tree. This species is not confined to the island for which it is named, but occurs also on the mainland. But -- it does not grow on the adjacent mainland around Perth, instead the closest occurrence is near Dongara, some 321 km further north!
Why does Rottnest Island have no gum trees? Why is the tea tree species so distant from its kin? These apparently trivial matters are a small part of a greater truth which we will try to uncover. For the moment we can be content with the important and perceptive observation of Somerville which is quoted at the head of this article.
The Spread of Plants
Plants and animals together have been of basic importance to man, extending back to the earliest times that a creature which could be called a man first existed. Together, they provided man with all of his food, most of his clothing, and much of his fuel and raw materials for artefacts and construction.So it is natural that he has had an intense interest in these two great divisions of life, kingdoms they are justly called. Through this interest and its resulting knowledge he has been able to exploit the twin kingdoms to his own advantage, and make himself the emperor over both.
Over the years of history, one topic which received attention was the way in which plants were able to disperse themselves, to spread and propagate themselves into new areas. Some of the mechanisms used by particular plants are strange indeed, and the rich diversity and range of these mechanisms is well known. A thorough treatment of the subject appears in Ridley's book "The dispersal of plants throughout the world." [69].
Ridley's book gives many examples of long journeys made by plant seeds, found washed up on distant coasts, taken from the feet of migrating birds, carried in the fur of nomadic animals, and so on. Of course most seed dispersion mechanisms strive to separate seeds as far as possible from the parent plant, and many mechanisms clearly have the potential capability to achieve very rapid spread of the species involved. If a dandelion seed can drift 500 metres in the wind, we might expect an initial planting of dandelions to extend over a circle 10 km wide within 10 years. Does such a spread usually occur?
In fact, the answer is usually 'No'. Actual rates of spread of plants (and of animals too) are enormously smaller than their potential rates as worked out from the mobility of one generation of offspring. It is clear that this must be so, otherwise the spectrum of living creatures found in one place would be more or less identical to that of somewhere else not too far away. In actual fact, it is not terribly unusual for the species list of a given area to differ by more than half its contents from that of another area only 100 km away, even when the areas are directly connected and superficially similar in nature.
Proposition 2A
Actual rates of spread of plants are usually much less than the
potential rates of spread implied by the dispersion mechanisms
operating for an individual seed
There are a number of reasons which can be suggested for this. There may be an obvious physical barrier to the spread, such as a stretch of water or a high mountain range. There may be a more subtle change in the environment, such as a switch to a different soil type with different nutrients or moisture retention.
But the most important reason is a factor which can be called Ecological Pressure. Different plants, and different animals too, do not use the whole spectrum of conditions in which they exist. Instead, they occupy what are called 'ecological niches'. A clear example is the epiphytic plants which grow high up on the trunks of tall trees in a tropical jungle. These plants are not parasites, they just have a strongly position-dependent ecological niche.
Other ecological niches are much more subtly defined. The 'boundaries' of the niche may be formed by such things as a change in soil texture or composition, proximity to water or rocks, exposure to winds moving in a particular direction, dependence on the presence of particular animals or other plants, and every possible permutation of factors like this.
In order to spread, a plant or animal species would usually need to displace some other creature which is already occupying the particular ecological niche for which it has evolved. In most cases, it is unable to overcome this ecological pressure, and so it does not succeed in spreading.
Proposition 2B
Plant and animal species do not expand their ranges easily, because
they are unable to overcome ecological pressure from other
species already occupying their ecological niches
There have been cases where ecological pressure is absent, and in these, rapid colonization and expansion may occur. A dramatic example occurred during the last century, when the explosion of the volcano Krakatoa removed the whole top of a small island in the sea near Java, wiping it completely clean. Within a few years, however, recolonization of the 'new' island recommenced, and now the plant cover is similar to that on adjoining islands.
That was a natural example. A more familiar example, based on the activities of man, is when areas of forest or jungle are cleared for the planting of field crops. The crops are planted, and shortly afterwards the weeds move in -- the clearing provided vacant ecological niches not only for the crops, but for the weeds as well. The human activity may be quite minor, such as digging a hole to plant a tree. It has been shown that weeds will invade trial plots in some open areas in Australia only if the soil is disturbed, adjacent plots with no soil disturbance are not affected. The minor action of disturbing the soil surface is sufficient to create a vacant ecological niche for the weeds. This does raise a neglected possibility for weed control.
Proposition 2C
Weeds may be controllable through manipulation of their
microecological surroundings, rather than through direct attack
by sprays or cultivation.
Plant Mutability and the Isocons
In this book I will be referring constantly to 'Isocons'. These are like the isobars on a weather map, the 'contour lines' showing areas of equal pressure, but instead of pressure they define the boundaries of an area of equal ecological conditions.In other words, isocons are lines drawn on a map which delineate the local boundaries of a plant's ecological niche. The area enclosed by an isocon is that which we would expect to be rapidly filled by a plant if there was no ecological pressure operating, say if disease had wiped out all its competitors for that niche.
Under settled conditions, the isocon for a given plant species is the same as its natural wild range. The point is that the plant has evolved to fit that range. We speak about 'the plant' as though it were a particular genetic identity, but in fact any plant species is a great mixture of different gene types.
The genes are continually mixing, recombining, and to some extent mutating, within a species. The boundary between one species and a closely-related one in the same family is really rather arbitrary, and taxonomists, the people whose work it is to define these boundaries, are continually arguing and moving them around to include or exclude particular plant populations.
Two related species share a high proportion of identical genes, for example it has been estimated that 98% of the genetic material of man and chimpanzee is the same. The cut-off really comes at the point where cross-compatibility is a factor.
If any individual in a given population can potentially interbreed with any other individual (of appropriate sex) in a second population, to produce fertile offspring, then those two populations are certainly in the same species. If there is no compatibility between any individuals of either population, the two populations belong to different species.
In practice these two states grade imperceptibly into one another, hence the fertile (!) ground for the taxonomists to argue on. It is not physically possible to test every individual of the two populations against each other, but what usually happens is that individuals at the ends of the ranges, those most widely separated, get more and more out of kilter.
Whatever, it is appropriate here to make the point that the ragbag of genes which we perceive to lie within the bounds of a given 'species' is in a state of continual flux, due to mixing, recombination, and mutation. A species today is different to the 'same' species a hundred, a thousand, a milllion years ago. It is as if an ant colony undertook a long migration, occupying a hundred years, across a continent. The individuals which reached the final point would all be different to those which started. And on a more basic level, the genetic constitution of the colony would have changed too, as characteristics favourable to the conditions met with were selected.
Proposition 2D
The total genetic constitution of a species is subject to
continual alteration, particularly if external conditions are
changing
Crossing the Isocons
Figure 2.1 shows the mapped distributions of some species of Canarium in the southeast Asian region. This is a largely tropical family of trees, which includes the Pili nut and the Java almond. It can be seen that some of the species occur over fairly wide areas, while others are small in range.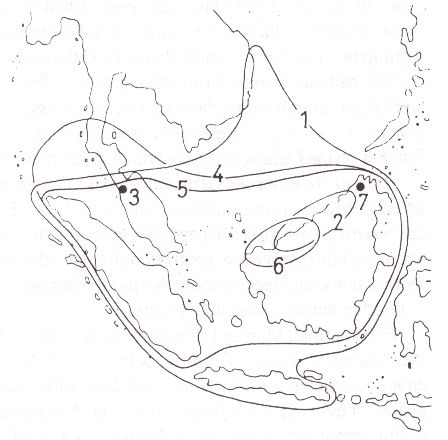
Fig. 2.1. Distribution of some species of Canarium [48]
In particular, note the black dot marked '7' on the map. That dot is the total range of the species Canarium kinabaluense, and it coincides exactly with the site of Mount Kinabalu, the highest mountain in southeast Asia. Due to this height, the physical conditions on the mountain are naturally quite different to those below the mountain -- in other words, the isocon for C. kinabaluense is dependent on altitude.
Another way of looking at this situation is to ask how this species got to be a separate species, how it evolved from some former common stock. The answer is that, in the past, a group of the genetic elements in the common stock which were particularly suited to the high-altitude conditions separated off to give the new species.
Perhaps a mutation occurred in some of the Canarium trees which were lapping up against the potential isocon ring round the mountain. Perhaps there was slow selection of cold-tolerant genes already present. It is even possible that the change was in the reverse direction, with the mountain species being the original one, with the others adapting from it as the climate warmed up or the sea-level fell.
In any event, to cross the isocon, the species had to undergo adaption and change. These changes were great enough to reach the level where the resulting plant population has been classed as a separate species.
This example of an isocon has a clear physical basis, it depends directly on altitude. Other isocons are more subtly based. For example, the different levels of a dense tropical jungle are worlds apart -- on the upper canopy there is intense light and may be burning heat and fierce winds, accompanied by large daily temperature changes. Down below, on the forest floor, it is dim and still, with much less variation.
In one site in the Central American jungles, it has been found that of four species of bee present there, two are confined entirely to a lower level, and two to an upper one -- there is no mixing [62]. Plant species may be similarly stratified, and these more subtle isocons lead to the situation where two distinct but related species evolve in what appears to be the same geographical area, because the isocon separating them is not based on location, but on some other factor.
Rates of Change in Species
Rottnest Island has one species of bird in the family Lichenostomus, the Singing Honeyeater, L. virescens. This species is the only one of the family on the island [70].The same species also occurs on the adjacent mainland, together with other species of honeyeater. What is interesting about the Rottnest population of Singing Honeyeater is the fact that it is appreciably different from the mainland population of the same species. The Rottnest birds tend to be larger, some 20% heavier, and much darker on the lower surface.
It has been suggested that these birds are evolving to fill the niches which are occupied by other honeyeater species on the mainland, because these other species are lacking on Rottnest. They are on the way to becoming a separate species.
It has been stated that physical isolation of the two populations occurred only about 7000 years ago -- 0.007 my, a tiny amount on the scale we have been using.
Now let us look at the situation in another famous set of islands, the Galapagos islands off the coast of South America. Back in September 1835, Charles Darwin visited the Galapagos in the ship the 'Beagle', and the observations that he made there [25] laid the foundations for the Theory of Evolution.
These islands are isolated some 900 km off the mainland of Ecuador, and consist of a group of about 5 larger islands and many small ones, all of recent volcanic appearance. What Darwin observed there can be summarized in terms of what are now called Darwin's Finches, a group of small birds found only on the Galapagos. He found that this group of birds, clearly related, had taken on differing characteristics from one island to another, and the differences were great enough so that the birds had to be classed under different species, and even different genera. A current classification [34] shows that there are now 13 species of Darwin's Finches on the Galapagos, all of which have apparently evolved from a single common stock.
It should be emphasized that most of the Galapagos islands are within sight of one another, and that these were normal birds, capable of flight. Even so, the forces which govern evolution are powerful enough so that even a relatively small separation in distance was enough to bring about genetic divergence, great enough to take the birds into different species and genera. And there was nothing special about this particular case of the finches, the same thing had happened with other animals, and with plants.
For example, David Murray has shown [54] that of the 543 native species of higher plants on the Galapagos, around 40% are endemic, that is, they are species found only in that local area. These endemic species must have evolved from older stocks which elsewhere have either disappeared or evolved in a different direction. Murray has calculated that "the minimum number of immigrant progenitors needed to account for indigenous angiosperms [higher seed plants] is 306". We will return to the point of immigration later, now only noting this as the assumed route of origin of the plant populations.
Of course when we move up to the next broader level above that of species, to the level of genus, the level of endemism is much less. The majority of the plant genera, such as Acacia (wattles), Psidium (guavas), and Opuntia (prickly pears) have species which are native elsewhere.
The Galapagos appear to be relatively young in geological terms, having existed in isolation for no more than 3 my , and possibly as little as 0.5 my [34, 54]. This period has been long enough to evolve 13 species of finch from the original one. On another isolated site, the Hawaiian archipelago, Grant has shown [34] that at least 42 species of honeycreeper finches have evolved from a single ancestor during the last 6 my.
From these two figures we can derive a rule of thumb to estimate the rate at which species will evolve and split in the absence of ecological pressure. If we assume that a species will diverge enough to split in two every one million years (doubling every my), it would take about 3.5 my to produce the 13 Galapagos finches and about 5.5 my to end up with the 42 Hawaiian finches -- both close enough for a first stab.
Proposition 2E
In the absence of ecological pressure, a species diverges into
two species roughly every million years
Changing Climates -- March of the Isocons
So far we have only looked at the situation as of now, with today's conditions of climate and elevation in particular parts of the world. In fact, we know that both climate and sea-levels have varied dramatically in the past in most parts of the world.We can also represent this position by saying that the isocon lines do not stay fixed with time, but move around as external conditions change. As the sea-level rises, the major sea/ land isocon moves 'inland' into what was previously higher ground, as the oceans fall it moves out to sea. As the climate cools, the major frost/no-frost isocon moves towards more equatorial latitudes, and as the climate warms, so it drops back toward the poles.
In this way, every species population is subjected to a sort of ecological massage. The isocon boundary bag within which it lives is pushed back and forth, up hill and down, north and south. Every species is a nomad.
This picture leads to an important conclusion. As the isocon boundary bags are moved around over the terrain by various external changes, it is inevitable that some will cease to exist. Some will fall over a cliff into the sea (as when an island flora is inundated); others will evaporate off the tops of mountain (as when warming causes the frost line to cease to exist). When this happens, either one population of a species, or even the whole species, will also cease to exist.
Proposition 2F
Species tend to die out when the ecological niches in which
they exist are eliminated, and this elimination is promoted by
continuing changes in external factors such as climate and sea-level
There is also a most important counter-conclusion. When subjected to this ecological massage, when the isocon bag is nearing the edge of the cliff, some species will have the genetic resources to adapt and cross the isocons, thus becoming a new species. Natural selection is known to promote the formation of new varieties and species, so changes in natural selection pressures must speed the process of evolution up even more.
Proposition 2G
Changes in external conditions increase rates of natural
selection and evolution
These two last propositions are clearly at the two ends of the tug-of-war rope. They imply that the rate of formation and extinction of species is much greater than has been assumed. During the last 2 my, there have been five major glacial advances and five retreats (five 'ice ages'). Most isocon bags will have been shifted some hundreds of kilometres during each of these ten trips. Those situated on small islands must have had a hard time!
Conclusions on Plant Evolution and Spread
We have built up a picture of previously largely-unrecognized rapid changes in the genetic bases of plant and animal species. It is clear that these species have the capacity to undergo marked changes in their total genetic bank over what are very short periods on the scale of the Earth's history.In fact this conclusion is obvious, when we look at the rapid changes in species which have been brought about by man, purely through simple processes of selection, in periods measured in decades, not even centuries. An important point here is that most of these changes are very visible ones -- bigger and brighter flowers or fruit, rapid or dwarfed growth, and so on.
Under natural selection, many of the genetic shifts which occur in species will not show up very visibly. If an isocon shift moves a plant population from a sandy soil onto a clay soil, selection of the population will occur to cope with the change in soil type and the associated nutritional factors which this implies. But there may be no visible change at all in the appearance of the plant, and even at the detailed genetic level, it could be hard to find a quantitative difference.
The ginkgo or maidenhair tree, a fascinating nut tree, has been claimed as the oldest 'living fossil'. Standing on the border between the major plant groups of gymnosperms (conifers) and angiosperms (broad-leaved trees), the ginkgo was first recognized in fossil leaf impressions dating back some 200 my, and was not originally known to Europeans as a living plant.
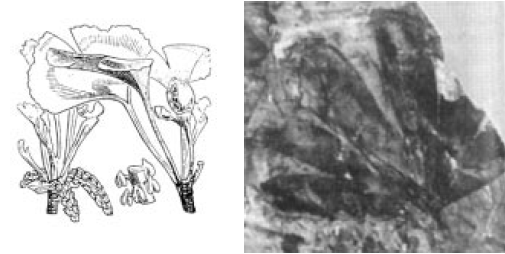 Fig. 2.2. Images of living and fossil ginkgos
Fig. 2.2. Images of living and fossil ginkgos
However in 1712 an employee of the East India Company, Englebrecht Kaempfer, noted some cultivated ginkgos in Japan [36]. In later European travels in China, a number of living gingko trees were found, mostly planted around temples. It is still uncertain whether there are any 'natural' or wild occurrences of ginkgos, although an area in Chekiang province in China has been suggested.
The point that is being made here is this. It has been claimed that the ginkgo has remained "unchanged for 200 million years". This is most unlikely. While modern ginkgos are undoubtedly related to those which produced the ancient fossil prints, they would certainly not be classed as the same species as the fossils if the latter were alive today -- the underlying genetic differences must be enormous. Later on we will see why this must be so.
In fact it is likely that no species is capable of maintaining its genetic identity for any very long period, say much more than around 10 my. Even less is it likely that this identity would be retained through a major physical turnaround like that at the end of the Mesozoic Era, some 70 my ago. Reasons for this major upheaval will also be looked at later in this book.
Proposition 2H
No species can maintain its genetic identity for long periods,
more than around ten million years
Are these suggestions reasonable in the light of logic and evidence? I have suggested that species are capable of splitting into two, doubling themselves, every million years (and this may turn out to be a low estimate). If there are a million different species of plant existing today (and current estimates approach this figure), then in the next 10 my there could be ten doublings, to produce around one thousand times as many species as now.
It is believed that the 'density' of species has increased appreciably since life appeared on Earth, with less diversity in the earlier years. This is a reasonable supposition anyway, as the complexity of particular species has reached higher levels, fitting them to operate more and more efficiently in smaller and more defined ecological niches -- the trend toward specialization.
Nonetheless, the actual increase in species density is far, far smaller than would be implied by general application of Proposition 2E, doubling every my. What is happening, of course, is that species are dying out as well as being formed, and these two processes are more or less in balance -- the situation is not too far from a steady state as far as total number of species is concerned. Just like the atoms of a radioactive substance, we can assume that species have a half-life, the average time needed for half the total number to disappear. The actual numbers depend directly on Proposition 2E and the assumption of a long-term steady state:
Proposition 2I
The half-life of a species is approximately one million years.
This last proposition is actually a modification of 2H before it; the 10 my in 2H becomes just the time at which the probability of a species surviving has become very small (around 0.1%).

(Full list of references at NURefs)
[25]. Charles Darwin. Journal of researches ... during the voyage of HMS Beagle. 2nd ed, 1860.
[34]. Peter R Grant. Ecology and evolution of Darwin's Finches. Princeton Univ. Press, 1986.
[36]. D A Griffiths. Ginkgo - tree of antiquity. West Australian Nut & Tree Crop Assn Yearbook/ 12 p62-70, 1987.
[48]. P W Leenhouts. Canarium. Blumea/ 9(2) p275-331, 1959.
[54]. David Murray [ed]. Seed dispersal. Academic Press, 1986.
[62]. Donald R Perry. The canopy of the tropical rain forest. Scientific American/ 251(5) p114-122, Nov 1984.
[69]. Henry N Ridley. The dispersal of plants throughout the world. Reeve, London, 1930.
[70]. Dennis Saunders & Perry de Rebeira. The birdlife of Rottnest Island. DAS & CPdeR, Western Australia, 1985.
[75]. W Somerville. Rottnest Island in history and legend. 4th ed. Rottnest Island Board, 1976.
NU003: Continental Drift And Earth Expansion
NU001: Setting The Scene
Version 2.0, 2004, PDFs etc on World Wide Web (http://www.aoi.com.au/matrix/Nuteeriat.htm)
Version 3.0, 2014 Sep 12, Reworked from Chapter 2 of "Nuteeriat" as one article in a suite on the World Wide Web.